by Ben Best
Viewed as a whole, the human nervous system is primarily concerned with the processing of sensory input or with the execution of motor output (physical action). Sensory input and motor output are relevant for both an internal and an external environment. For example, sensory input from the internal environment provides a person with an awareness of thirst. Sensory input from the external environment provides that person with information on how to obtain water. Motor activity directed to the external environment can result in the ingestion of water. Motor activity in the internal environment can assist the ingestion of water until sensory information from the internal environment provides evidence that enough water has been consumed.
The internal environment is primarily the domain of the autonomic nervous system (ANS), a system that is (for the most part) not subject to conscious awareness or control. The kidneys, intestines, lungs, glands, blood vessels, heart muscle, etc. are under the control of the ANS. The two divisions of the autonomic nervous system are the sympathetic ("fight or flight") and the parasympathetic ("rest and digest"). Whereas sympathetic stimulation increases heart rate, constricts blood vessels and decreases peristalsis in the intestine, parasympathetic stimulation has the opposite effects.
Anatomically, the human nervous system is divided into two portions: the central nervous system (CNS) and the peripheral nervous system (PNS). The autonomic nervous system is a functional division of the nervous system which is part of both the CNS and the PNS.
|
|
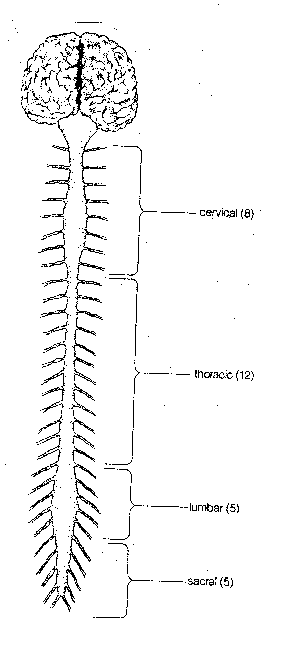
The central nervous system (CNS), consists of the spinal
cord and the brain. The spinal cord is housed in the backbone, with 30
pairs of nerves receiving sensory information and sending motor information
to the body. The spinal cord and its nerves are divided into four regions:
cervical (neck), thoracic (chest), lumbar (loin) and sacral
(pelvis). The
sensory information is mostly somatosensory: touch, temperature, pressure
and pain ("somato" = "body"). Axons in these nerves tend to cross to the
opposite side of the spinal cord so that
sensation-in and control-of the
right side of the body are in the left brain, and vice versa. Within
the spinal cord axons ascend-from and descend-to the brain in well-segregated
tracts: for crude touch, for pain&temperature, for motor control,
and for proprioception&kinesthesia (muscle, tendon & joint position as
well as muscle tension and movement information).
|
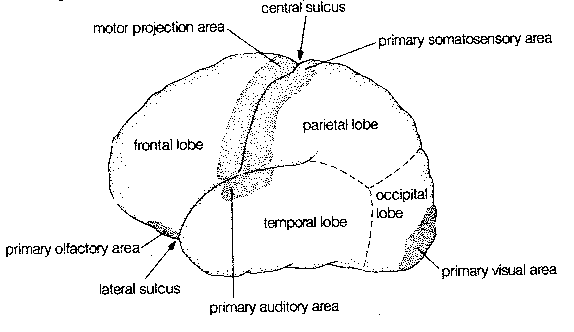
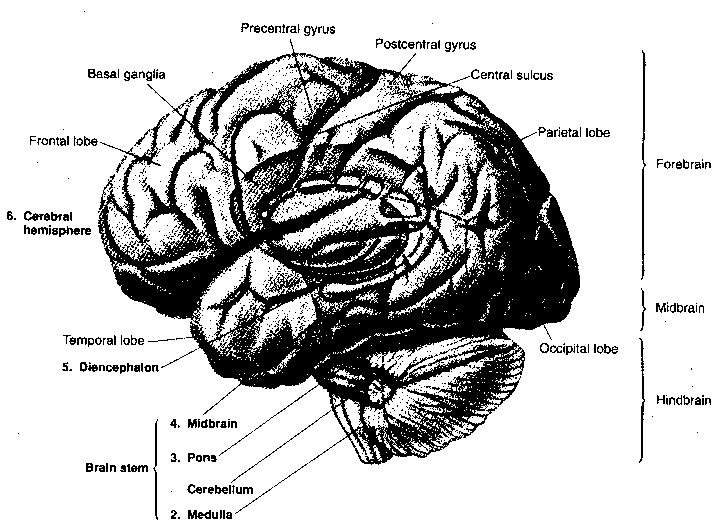
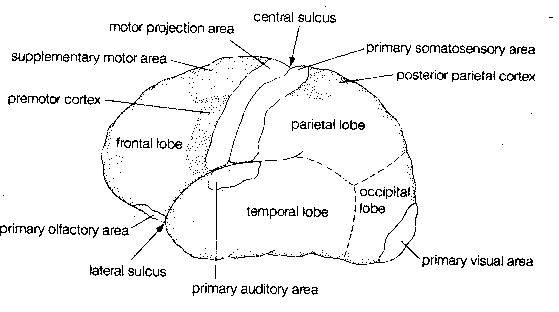
Crowning the CNS is the cerebral cortex, which is often regarded as the seat of the mind. The cerebral cortex has the appearance of a ball of worms. A "worm" (elevated ridge) is called a gyrus, whereas a "valley" between worms is called as sulcus.
The exterior of the cerebral cortex is divided into four lobes: frontal, parietal, temporal and occipital. Much of the occipital lobe is devoted to processing sensory input from the eyes whereas a portion of the temporal lobe is devoted to processing sensory input from the ears, and a portion of the frontal lobe is devoted to processing sensory information from the nose. The frontmost portion of the parietal lobe (the postcentral gyrus) is concerned with representing sensory information from the body — somatosensory information). The rearmost portion of the frontal lobe (the precentral gyrus) is concerned with conveying motor information (initiation of voluntary movement) to the body,ie, muscle control. The central sulcus separates the precentral gyrus from the postcentral gyrus.
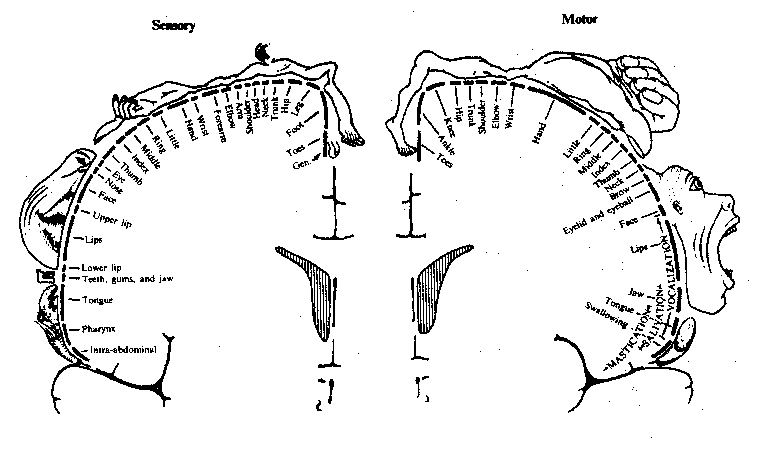
Precise localization of the body parts supplied by both the
somatosensory and somatomotor gyrus of the cerebral
cortex is possible. Parts of the body that have acute sensation or fine
motor control, such as the face and hands, occupy a disproportionately
large area of the cortex.
|
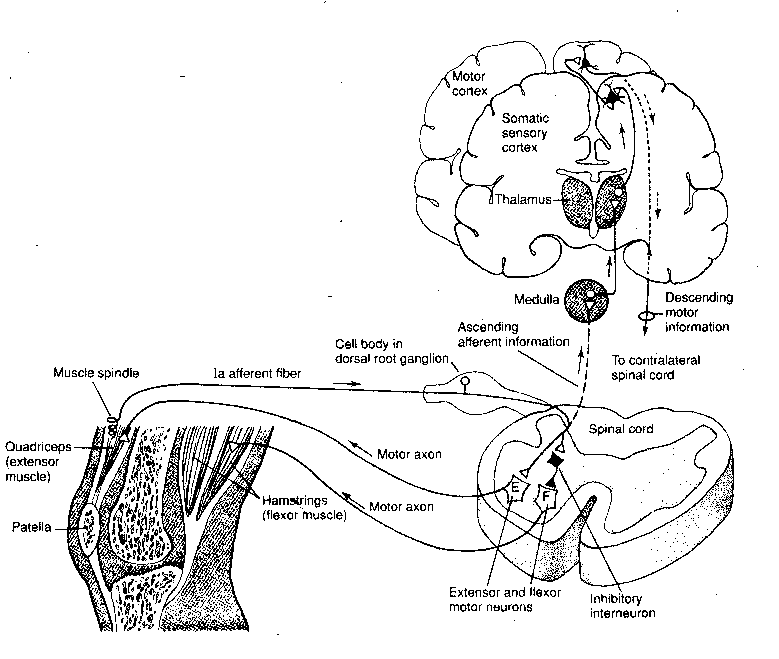
The simplest form of movement in response to sensation is
probably the stretch reflex. For example, in the knee jerk reflex,
tapping on the knee pulls on the tendon of the extensor muscle that
extends the lower leg. The stretch of the muscle in response to the
pull of the tendon results in a signal to the spinal cord along the afferent
fiber connected to the muscle.
The afferent fiber from the extensor muscle spindle makes a direct connection
to a motor neuron leading straight back to the extensor muscle. The same
afferent fiber also sends a branch to an
inhibitory interneuron that repetitively fires inhibitory impulses to
the flexor muscle on the back of the thigh, causing it to
remain flaccid. Inhibitory and stimulatory axons in the
pathways from the brain could increase the complexity of responses to
stimulation.
|
Intelligence, memory and emotion are perhaps nothing more than vast arrays of interneurons which evolution has interposed between sensory stimulation and motor response. Touching a hot stove, for example, results in rapid and forceful withdrawal of the arm. However, a sufficient act of will-power can suppress this reflex by sending enough inhibitory impulses along descending fiber tracts from the somatomotor region of the cerebral cortex.
Locomotion is a voluntary action with a high degree of automaticity. Complex spinal circuits act as central pattern generators which can control walking, trotting and galloping. A cat whose descending spinal fibers have been severed can still walk when placed on a treadmill. Moreover, increasing treadmill speed can change the locomotive pattern from walking to trotting to galloping. In walking, the left and right hindlimbs are exactly out of phase, whereas in galloping the hindlimbs are exactly in phase (they flex and extend together).
Circuits controlling movement in the cerebral cortex can also be programmed, as should be evident from the apparent automaticity of movement often demonstrated in the driving of an automobile.
|
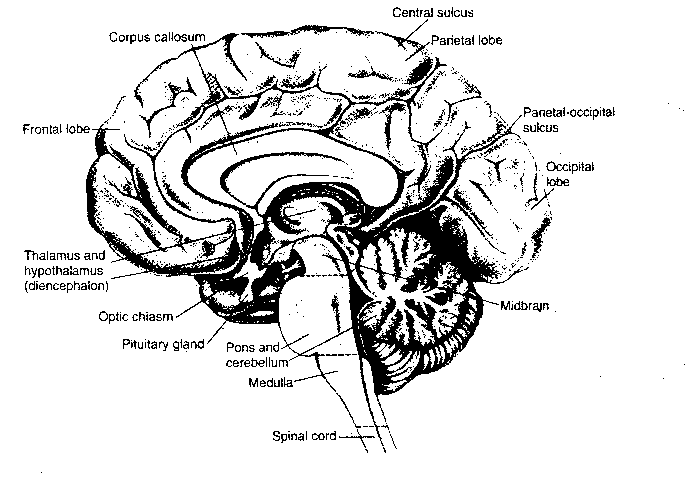
Above the spinal cord is the brain, composed of the brain stem (medulla oblongata, pons and midbrain), the cerebellum, the diencephalon (thalamus and hypothalamus) and the cerebral hemispheres (basal ganglia, amygdala, hippocampus and cerebral cortex).
The medulla, the pons and the midbrain are a continuation of the spinal cord into the brain, hence they are the "brain stem". Sensory and motor axons controlling the head, mouth and face connect to the brain stem by cranial nerves. Also among the cranial nerves are the nerves for the special senses of vision, balance, hearing, taste and smell. Finally, cranial nerves carry parasympathetic innervation of the autonomic ganglia that control breathing, heart rate, blood pressure, coughing and swallowing. An important distinction between the brain stem and the spinal cord is the fact that the brainstem contains many nuclei,ie, clusters of neurons organized into functional groups. In addition to the discrete nuclei of the brain stem, there are small groups of diffusely distributed neurons making-up what is known as the reticular formation (the arousal center of the brain).
|
Half of the cranial nerves enter the brain at the level of the medulla oblongata. Only one of these nerves is associated with a special sense, and it is the special sense which is the least a part of conscious awareness: the sense of balance. Four medullary nuclei are concerned with balance — the vestibular nuclei. The medulla controls a number of vital autonomic functions, including digestion, breathing and heart rate. Somatosensory axons in the dorsal column of the spinal cord synapse on neurons of the dorsal column nuclei in the medulla, and somatosensory information then projects upward to the thalamus as a tract of fibers known as the medial lemniscus.
Wrapped around the back of the brain stem is the cerebellum (Latin
for "little brain"). Although the cerebellum constitutes only 10% of the brain
by volume, it contains half of all the neurons in the brain. Analogous in
appearance to the cerebrum, the cerebellum consists of an outer mantle of gray
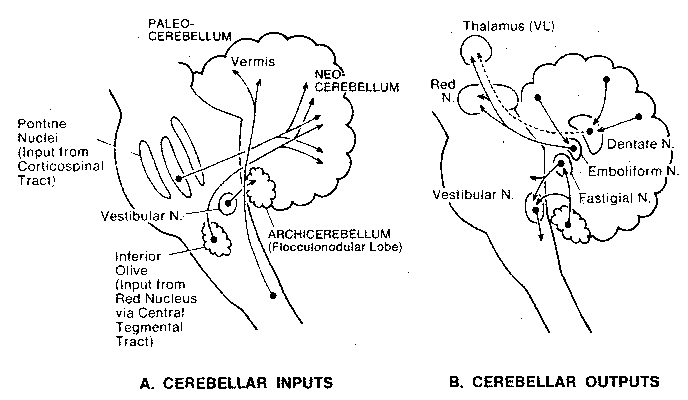
matter, internal white matter, and three deep nuclei: the fastigal, the
interposed and the dentate nuclei. Input to the cerebellum is
from all levels of the central nervous system to the cerebellar cortex,
but output from the cerebellar cortex is via the deep nuclei.
|
The main function of the cerebellum is the coordination of motor activities. Commands to initiate movement come from the somatomotor cerebral cortex, but the cerebellum fine-tunes motor control, giving smoothness of motion and exactness of positioning. The cerebellum makes comparisons of internal and external feedback signals to correct ongoing movements. Lesions of the cerebellum result in movements that are jerky and that overshoot or undershoot their intended mark. The cerebellum is the site of much motor learning — in particular, the motor learning of well-timed and co-ordinated movements and sequences.
Inputs to the cerebellum come from mossy fibers originating
in the pontine nuclei and other brainstem sites, and climbing
fibers from the inferior olivary nucleus of the medulla. Climbing
fibers synapse strongly on Purkinje cells, which are the only
neurons producing output from the cerebellar cortex. Purkinje cell
axons are all inhibitory (using GABA transmitter) and all terminate
on deep nuclei cells. Purkinje cell activity is modulated by a variety
of inhibitory interneurons of the cerebral cortex, which receive diffuse
stimulation from fibers originating in the raphe nuclei and
locus ceruleus nuclei of the brain stem.
|
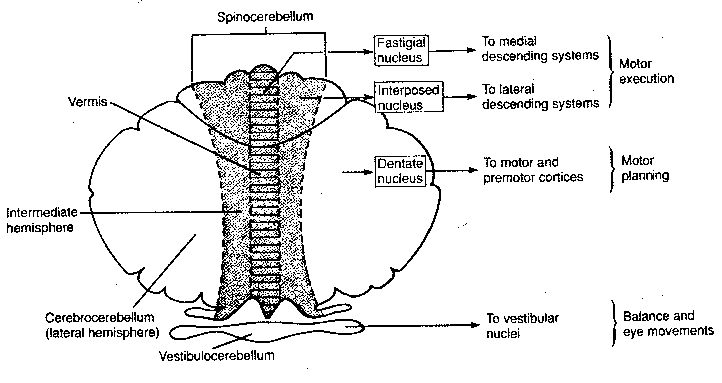
The cerebellum consists of three functional regions: the vestibulocerebellum, the spinocerebellum and the cerebrocerebellum. The vestibulocerebellum (archicerebellum) was the first to appear in evolution, and it controls eye movements and balance through its connections with the vestibular nuclei of the medulla. Most of the input to the spinocerebellum (paleocerebellum) is somatosensory, and through its outputs it controls the execution of movement and regulation of muscle tone. The cerebrocerebellum (neocerebellum) communicates with the cerebral cortex through the pontine nuclei of the pons. In fact, the major function of the pons is the mediation of information about movement between the cerebrum and the cerebellum — as part of movement planning. Portions of both spinocerebellar and cerebrocerebellar output go to the red nucleus in the midbrain, which integrates information about distal motor control (such as the hands and fingers). (The arms, hands and fingers are the primary means of human manipulation of the environment, as distinct from locomotion in the environment. Direct tracts of axons from the cerebral cortex to these appendages are their primary means of control.)
The tectum in the midbrain is the key structure in nonmammalian vertebrates for integrating sensory inputs and for controlling motor outputs. But in higher mammals, the tectum's function is more narrowly restricted to co-ordinating head and eye movements, and to mediating visual reflexes concerned with pupil dilation and eye movement. The tectum is composed of an inferior colliculus (mediating sound) and a superior colliculus. The superior colliculus coordinates visual, somatic and auditory information in adjusting movements of the head and eyes toward a stimulus.
|
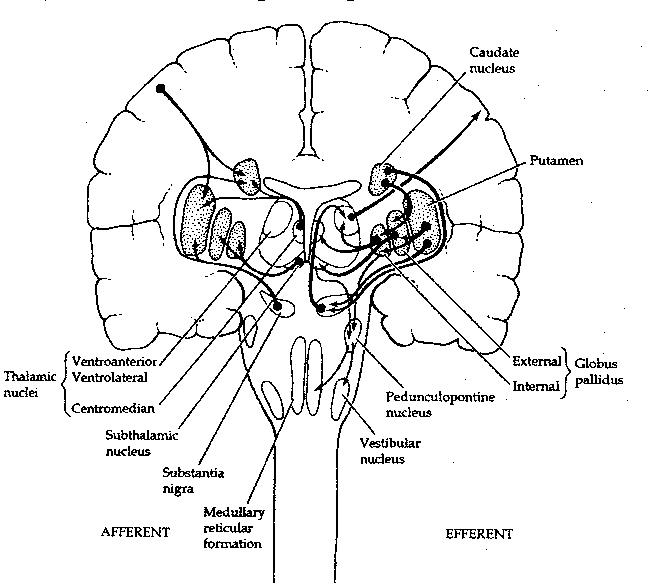
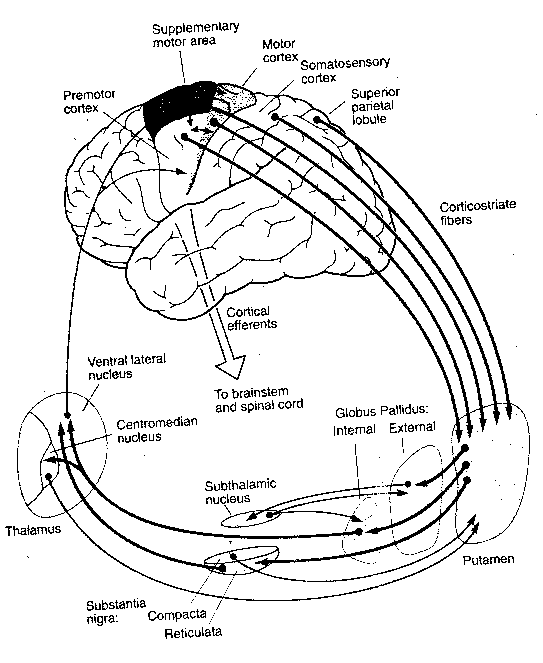
The basal ganglia are so-called because they lie mostly within
the base of the cerebral hemispheres.
Literally speaking, however, two of the
functional units of these large nuclei lie in the midbrain: the substantia
nigra and the subthalamic nucleus. The caudate nucleus is actually
just an extension of the putamen and those two structures are referred
to collectively as the striatum. Essentially, the basal ganglia receive
inputs from the cerebral cortex to the striatum, the inputs go onward to
the globus pallidus and then to the substantia nigra, which sends signals
back to the cerebral cortex via the thalamus. What could be the purpose of
this loop? Evidently the basal ganglia function to maintain the muscle tone
needed to stabilize joint position (as, for example, holding a glass of
water while talking) or to inhibit muscle tone during the initiation of
movement. Interruption of the feedback loops of the basal ganglia by damage
to one of its structures results in the uncontrollable oscillations manifested
as tremors.
|
Anatomically, the thalamus is divided into three groups of nuclei: the anterior, the medial and the lateral nuclear groups. The lateral nuclear group has the greatest number of subdivisions, which can be grossly divided into the ventral tier of nuclei, the dorsal nuclei and the large posterior pulvinar. The thalamus is a relay station for signals to the cerebral cortex. Although some motor information passes through the thalamus, most of the input to the thalamic nuclei is sensory.
Auditory information from the ears, for example, travels along nerves to the lateral superior olive and the medial superior olive of the medulla (the latter has cells which can detect phase differences between a sound reaching the two ears, allowing a person to localize the source of the sound in space). From the olive, auditory axons travel upward as a bundle (the lateral lemniscus) to the inferior colliculus in the midbrain tectum, and then upward to the medial geniculate nucleus in the thalamus, which projects axons onto the primary auditory cerebral cortex.
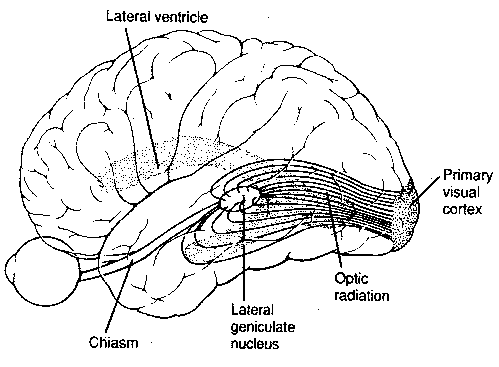
By contrast, the optic nerve from the eye sends a small branch of axons to the superior colliculus of the midbrain, while most of the axons terminate in the lateral geniculate nucleus of the thalamus, which projects axons onto the primary visual cerebral cortex.
Somatic information from axons in the medial lemniscus synapse on the ventral posterior lateral nucleus of the thalamus before being relayed to the somatosensory cortex. The medial dorsal nucleus of the thalamus relays olfactory (smell) information to the cortex, and the ventral posterior medial nucleus of the thalamus relays gustatory (taste) information. Inputs from the cerebral cortex and the reticular formation to the thalamus are believed to exert some control over what sensory information the cerebral cortex receives.
If the purpose of this essay is to find the mind in the brain, it would seem thus far that the quarry has eluded us. It seems reasonable to suspect that the mind may reside in the structures not yet covered: the cerebral cortex, the limbic system and perhaps in the hypothalamus. If this is true, the argument for "neuropreservation" can perhaps be carried further than it has been. For it might now seem that only the latter brain structures are necessary for the preservation of identity.
But what if most of the latter brain structures seem every bit as mechanical as the ones we have covered? Would that mean that we would only need to preserve some tiny portion of the brain to retain identity? A deterministic view would cause us to predict that all of the structures we have not yet covered are are mechanical as the ones we have covered. In that case, mechanistic neural function could not be a criterion for disposability, since we still do not regard ourselves as disposable, even if we are mechanical.
If learning and memory are to be the criteria for personal identity preservation, does that include motor learning? Is the ability of my cerebellum and basal ganglia to assist me in putting a basketball through a hoop — or catch a baseball — essential to my being? I am conscious of my ability to localize the origin of a sound, but that ability does not seem to be a part of my consciousness any more than is the ability of my heart to pump blood. To be sure, my consciousness leaves its imprint in my body much as a foot leaves an imprint in the sand. It may be that the footprint is worth preserving because of the information it provides about the foot.
|
The analysis of vision is almost an obsession with neurophysiologists,
and what they have learned can teach many lessons about the operation of the
brain/mind. Fibers from the lateral geniculate nucleus in the thalamus
project onto the primary visual cortex (occipital cortex region) like a
projector projecting an image on a screen. In one experiment a monkey was
trained to stare at a pattern that looked like the bullseye of a dartboard —
concentric circles with spokes from the center. The monkey was injected
with radioactive glucose which was absorbed by brain neurons most in
use. When the animal was sacrificed and its occipital cortex was
observed, the "bullseye-with-spokes" pattern was clearly visible on the cortex.
Thus, a map of the field of view exists in the primary visual area of the
occipital cortex.
|
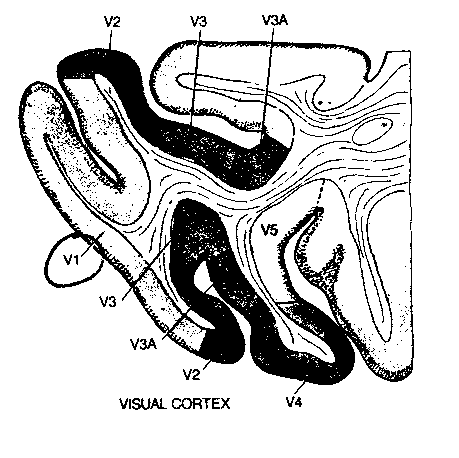
Close to the primary visual area (V1) of the cortex are cortical
areas composed of "columns" of neurons that are designated V2,V3,V4 and V5.
Neurons in area V3 are primarily sensitive to form, those in area V4 to
color and those in V5 to motion. This typifies what is seen in
studying the function of neurons in the cerebral cortex: "ensembles" of neurons
act as functional units for the processing of specific types of information.
|
Processing of visual information in the cortex is passed from
one ensemble of neurons to the next, to achieve higher levels of
abstraction: from sensation to perception to recognition. Objects
with very different shapes can be immediately recognized (identified) as
being cats, thanks to automatic processing of neuron ensembles. Information
form the Occipital Cortex ("OC") projects down the temporal lobe (toward
"TE") for object recognition ("what systems") and upward
to the parietal lobe (toward "PG") for object location in space
("where" systems). (It may
be no accident that the "highest" visual association neurons — of the
| Visual Cortex Paths | Optic Nerves |
|---|---|
![[ Visual Cortex Paths ]](./FigII14.gif)
|
![[ Optic Nerves ]](./FigII15.gif)
|
|
Analogous to the visual cortex is the olfactory and auditory cortex, which have a primary sensation area and proximal areas for sound or smell recognition. Moreover, it is now clear that higher-level ensembles of neurons associated with motor activity can be found adjacent to the somatomotor cortex — in particular, the supplementary motor area (SMA). In one experiment using PET (Positron Emission Tomography) scanning to monitor neural metabolic activity, human subjects asked to move their fingers demonstrated increased metabolic activity in both the SMA and the motor cortex. When the subjects were asked to mentally rehearse the finger movements without actually moving them, SMA neurons were metabolically active, but motor cortex neurons were not. The SMA, therefore, contains ensembles of neurons dedicated to the planning of movement.
|
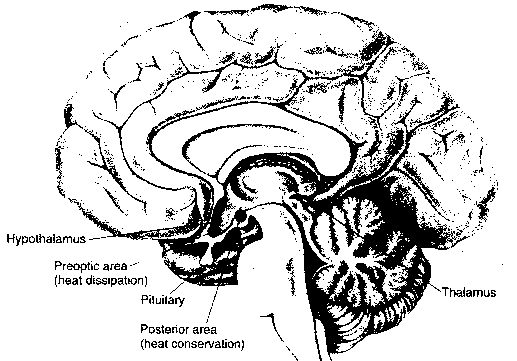
The hypothalamus is primarily concerned with regulation of the
internal environment of the body. It controls the pituitary gland, which
is the master gland of hormone regulation. The hypothalamus also is a center
for sensory and motor activity of the autonomic nervous system. Finally, the
hypothalamus is a center of motivation.
|
The hypothalamus controls temperature regulation in the body. Neurons in the hypothalalmus are responsive to both local warming and skin temperature. Electrical stimulation of the anterior hypothalamus causes dilation of blood vessels and suppression of shivering — resulting in a drop in body temperature. Electrical stimulation of the posterior hypothalamus has the opposite effect. Long-term exposure to cold results in the release of thyroxine hormone, which raises body heat by increasing metabolic rate. Various disease states can result in the macrophage product interleukin-1 which enters the hypothalamus and induces fever.
Similar to the temperature set-point (target temperature), the hypothalamus has a set-point for weight which determines how much an animal eats. Another set-point regulates water intake, and the hypothalamus can either cause the experience of thirst or cause the release of vasopressin hormone (which reduces urine volume).
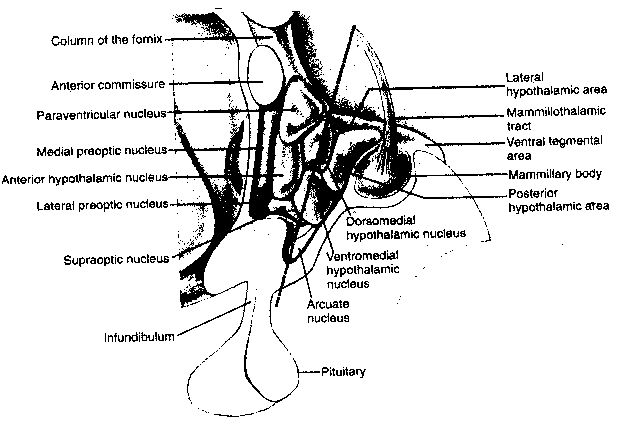
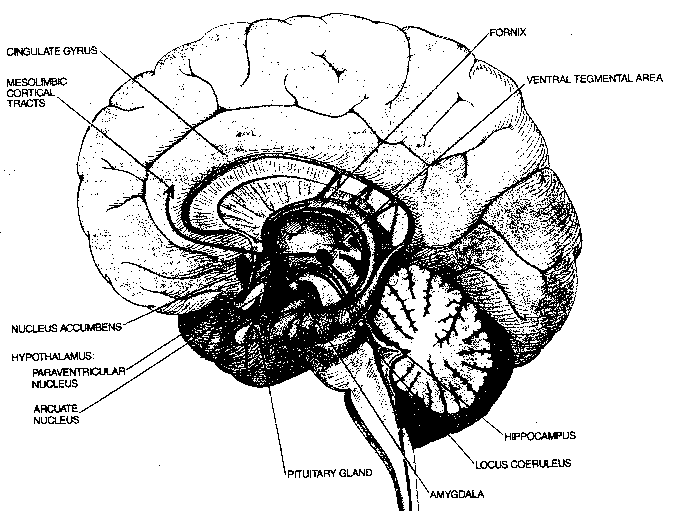
In the early 1950s it was discovered that bilateral lesions to the lateral hypothalamic area (LHA) will cause a rat to stop eating and drinking. A few years later it was discovered that rats that are enabled to electrically stimulate the LHA by pressing a lever will choose the electrical stimulation over eating, water or sexual activity — even to the point of death by starvation. Axons containing noradrenalin pass through the self-stimulation zone and if cocaine or amphetamine is used to encourage synaptic release of noradrenalin, self-stimulation increases. Tranquilizers which inhibit the action of dopamine suppress self-stimulation. It thus appears that the LHA is an important pleasure or reward center in the brain. Human neurosurgical patients who receive electrical stimulation of the LHA report feelings of pleasure and eurphoria. Nonetheless, it is evident that there are many other reinforcement sites in the human brain, for example, the nucleus accumbens of the limbic system. Cocaine is reputed to produce eurphoria primarily through enhancing the action of dopamine in the nucleus accumbens.
|
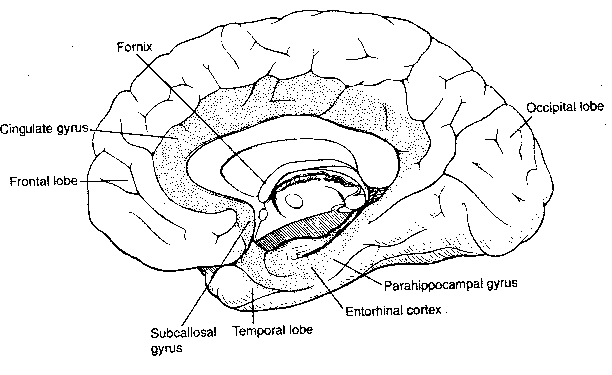
In addition to the four exterior cerebral cortex lobes — frontal, parietal, temporal and occipital — beneath the surface lies the most ancient area of the cerebral cortex: the limbic lobe. The limbic lobe includes the parahippocampal gyrus, the cinguate gyrus and the subcallosal gyrus. The limbic lobe communicates with the rest of the cortex primarily through the entorhinal cortex. The limbic system includes not only the limbic lobe, but the hippocampal formation, the nucleus accumbens and the amygdala. Some researchers even include the hypothalamus because it is so closely associated, anatomically and functionally.
The limbic system, in association with the hypothalamus, is regarded
to be the primary center of emotion. The association of the hypothalamus with
emotion should not be surprising, insofar as it controls the autonomic and
hormonal systems. Almost all emotions are associated with a response of body
organs — muscle constriction, heart rate alterations, etc. In fact, the
19th century American psychologist William James defined emotion to be
a "visceral response". In many ways, the experience of emotion is a
response of the central nervous system to changes in the body induced by
the autonomic system.
|
Groups of cells can be found in both the amygdala and the hypothalamus which respond to blood testosterone. Destructive lesions in the amygdala have given rise to "indiscriminate hypersexuality", and the stimulation of the amygdala can either induce or inhibit erections. Synchronous self-sustaining discharges are seen for neurons of the amygdala and thalamic nuclei during orgasm. (In the body, erections are the product of the parasympathetic nervous system, whereas orgasms are more the product of the sympathetic nervous system.)
The disease of rabies, which affects the hippocampus, can cause paroxysms of rage and terror. Stimulation of some portions of the amygdala produce rage reactions in animals, whereas the stimulation of other portions produce reactions interpreted as fear.
Insofar as the hippocampus is an important center of learning in the brain, its proximity to so many centers of emotion has interesting implications. For one, personal relevance of information determines what a person remembers, and this implies emotional involvement in the information. For another, learning is known to be facilitated by small amounts of anxiety and hampered by large amounts of anxiety.
|
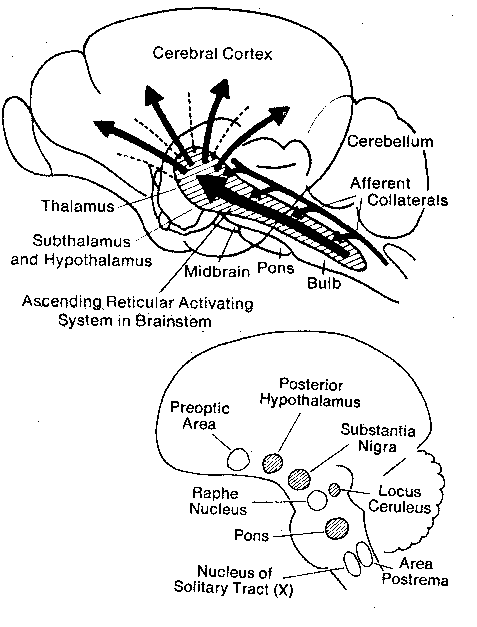
Located in the brainstem are an organization of nuclei and axon
tracts known collectively as the reticular activating system. Central
to this system are the reticular formation (distributed through the
medulla, pons and brainstem), the locus ceruleus, the raphe nuclei
and the preoptic region. The pontine reticular formation, the locus
ceruleus, the substantia nigra and areas in the posterior hypothalamus are
arousal centers,
whereas the nucleus raphe and preoptic area lead to
sleep. The reticular formation has four
functions:
behavioral
arousal, regulation of
muscular reflexes, coordination of autonomic
functions (like breathing and heartbeat) and the
modulation of pain sensations.
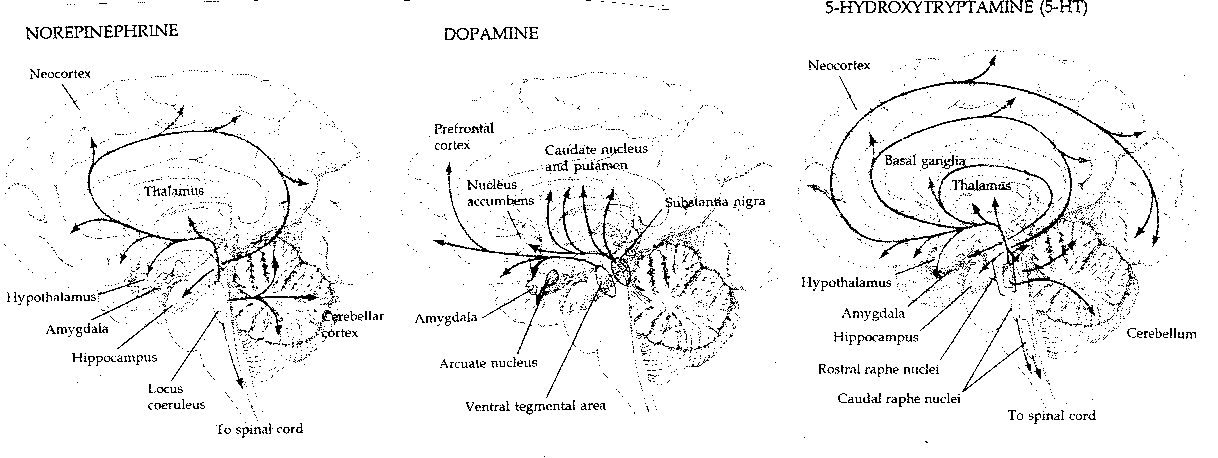
Half of all the neurons in the brain which use noradrenaline
(norepinephrine) as the
neurotransmitter are in the locus ceruleus. Four times as many neurons in the
brain use dopamine as opposed to noradrenaline as a neurotransmitter, and
these originate primarily from two midbrain nuclei: the substantia nigra
(a system related
to the disorders of Parkinson's Disease) and the
ventral tegmentum (the site of action of amphetamines and the
antipsychotic drugs). The number of brain neurons using serotonin
(5-HydroxyTryptamine)
is larger than the number using dopamine. Destruction of the serotonergic
raphe nuclei in cats eliminates their ability to sleep.
|
Many of the neurons in the reticular activating system spontaneously generate impulses as an inherent cellular property. Claims that perfect suspended animation of the brain would result in permanent quieting of brain electrical circuits ignores the inherent activating capacity of these neurons — they would resume activity upon restoration of functionality.
Electroencephalogram (EEG) rhythms of the cerebral cortex are believed to originate from the reticular nucleus in the thalamus, which (in turn) receives inputs from the brainstem reticular system, thalamocortical relay neurons and cortical pyramidal neurons.
An EEG is a recording of extracellular current flow associated with large ensembles of pyramidal neurons located in a section of the cerebral cortex just below the scalp. Because pyramidal cells are oriented parallel to each other, synchronized discharge leads to synchronized extracellular current. Studies of EEG activity are concerned with both amplitude and frequency (with frequency specified as cycles per second, or Hertz =Hz). In decreasing frequency, EEGs can be described as beta waves (13-30 Hz), alpha waves (8-13 Hz), theta waves (4-7 Hz) or delta waves (0.5-4 Hz). Alpha waves correspond to "relaxed wakefulness", and are typically observed over the parietal and occipital lobes. Beta waves are generally seen over the frontal regions during intense mental activity, and these waves have very small amplitude. Delta and theta waves are associated with deep sleep. Theta waves are associated with light sleep. For young adults, 20-25% of sleep time is spent in a state known as REM (Rapid Eye Movement) sleep, during which dreaming is most likely to occur. Although most of the cerebral cortex shows no EEG pattern during REM sleep (ie, cortical activity is desynchronized), the hippocampus shows highly synchronized theta waves during REM sleep.
An abnormally high level of synchronized discharge of large numbers of neurons results in a condition known as epilepsy. Epilepsy can result in jerking movements and/or loss of awareness (petit mal) or can be more intense, resulting in convulsions and loss of consciousness (grand mal). The hippocampus has the lowest seizure threshold of all cortical regions, and a loss of neurons in the hippocampus is commonly seen with epileptics. Epilepsy affecting primarily the hippocampus and amygdala is termed psychomotor epilepsy. In psychomotor epilepsy, patients often experience repetitive disturbing thoughts, mood alterations, a sense of impending disaster, depersonalization and/or inappropriate familiarity or unfamiliarity (deja vu or jamais vu).
![[GO TO BEN BEST'S HOME PAGE]](../../homeback.gif) HOME PAGE
HOME PAGE