by Ben Best
The sense of balance does not have connections in the cerebral cortex, but has terminal connections in the vestibular nuclear complex of the medulla and in the cerebellum. The senses of touch and taste are similar to vision in that the thalamus acts as the primary relay-station leading to the cortex. Nerve impulses from hearing, however, connect to nuclei in the medulla and then in the inferior colliculus of the midbrain before going through the thalamus to the cortex.
The snail-shaped cochlea of the inner ear is sensitive to both
frequency (pitch) and amplitude (loudness) of sound. Low frequency sounds
activate the basilar membrane at the apex of the cochlea, whereas
high frequency sounds have a greater effect at the basilar membrane at the
cochlear base. Loudness is detected both by the rate of nerve impulses and
by signals from certain cells that do not become activated until the
basilar membrane is vibrating at a high intensity.
|
|
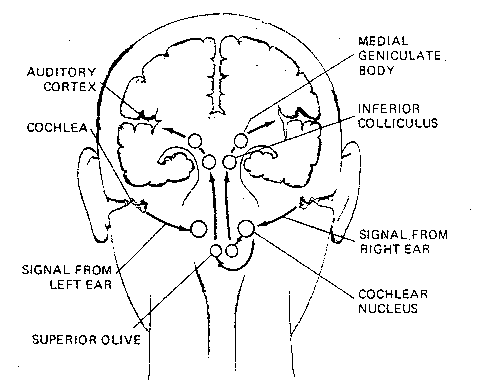
Nerves from the cochlea connect to
the cochlear nuclei in the upper medulla,
through the trapezoid body to the
superior olivary nuclei. The medial superior
olive is concerned with the localization
of sound in space based on time differences
of signals from the two ears. Although most
nerve fibers pass directly from the olivary
nuclei to the inferior colliculus as a tract
of fibers known as the lateral lemniscus,
many collaterals pass directly into the
reticular activating system. By this
means the entire brain and spinal cord can
be instantaneously activated by a loud sound.
|
Nerve fibers from the inferior colliculus
of the midbrain synapse in the medial
geniculate body (nucleus) of the thalamus. Fibers from the
medial geniculate nucleus of the thalamus terminate in layer 4 of the
primary auditory cortex (Brodmann area 41). Layer 5 projects back to the
medial geniculate nucleus and layer 6 projects back to the inferior
colliculus. Cells in the anterior portion of area 41 are more responsive
to low frequency sound, whereas cells in the posterior
portion are more
responsive to low frequency sound. Analogous to areas V1 to V6 in the
visual cortex (corresponding to color, motion, orientation, etc.) there are
multiple tonological maps in the auditory cortex, but these are nowhere
nearly as well understood as the visual cortex. It may be that these areas
associate loudness, distance or direction with tone. Brodmann area 42 is
the auditory association cortex which probably includes among its functions
the discrimination of sequences of sound patterns.
![[olfactory bulb and epithelium]](./FigVI3.gif)
Whereas vision is composed of 3 primary
sensations of color, there may be 50 or more
primary sensations of odor. This wide variety
of sensory response is mediated by odorant
binding proteins on the cilia of the olfactory
cells of the olfactory epithelium. Olfactory
receptors lose about half their responsiveness
within the first second of stimulation, and
very slowly thereafter. The well-known
adaptation to odors which is observed within
the first minute of exposure to an odor is undoubtedly mediated by
the brain, rather than by the receptors.
![[olfactory cortex]](./FigVI4.gif)
The olfactory bulb of the first cranial
nerve has two pathways to the cerebral
cortex. The oldest (in an evolutionary
sense) pathway leads to the limbic system
and, in fact, much of the limbic system
probably evolved from this olfactory
system. The primary olfactory cortex
is also known as the pyriform
(pear-shaped) cortex, corresponding to
the anterior portion of the uncus
of the parahippocampal gyrus of the
temporal lobe.
![[olfactory pathways]](./FigVI5.gif)
The lateral olfactory
tract connects directly to the uncus
of the temporal lobe of the cerebral
cortex without passing through the
thalamus. Fibers from the pyriform
cortex project to the entorhinal cortex
(Brodmann area 28), which contains an
olfactory association area that
sends projections to the
hippocampus. This portion of the
limbic system is undoubtedly
associated with the learning
of likes&dislikes of foods,
including the aversions that
occur with foods that have
been associated with
nausea&vomiting. A newer
olfactory pathway passes
through the medial
dorsal nucleus of the
thalamus to an area in
the orbitofrontal region
of the cerebral cortex.
![[anteriolateral touch sensation system]](./FigVI6.gif)
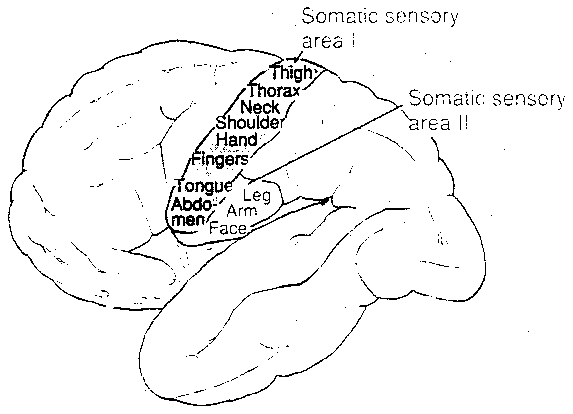
Touch sensation can be divided into two systems that take entirely different pathways from skin, muscles & joints into the central nervous system. The anteriolateral system (shown in the diagram) carries weakly localized sensations of temperature, pain, itching, etc. along two tracts in the spine, one anterior and one lateral. Most of these nerve fibers terminate in the reticular formation, the midbrain and 3 nuclei of the thalamus. Only a few fibers from one of these nuclei projects to the primary somatosensory area of the cortex, those from the ventral posterior lateral nucleus. The intralaminar nuclei project to the basal ganglia and diffusely to a wide range of areas of the cortex. The posterior nuclei project to regions of the parietal lobe outside the primary somatosensory area.
Most touch sensations which reach
the cortex travel as the medial
lemniscus (tracts not shown).
Nerve fibers in the medial lemniscus
are larger, and signals travel 2-3
times faster than those in the
anteriolateral system. Those touch
sensations provide information about
pressure, vibration and two-point
discrimination. These tracts terminate
on the gracile & cuneate nuclei
of the medulla. The medial lemniscus
continues upward, is joined at
the pons with fibers containing
facial information from the
trigeminal nerve, and terminate
in the ventral posterior nucleus
of the thalamus. Fibers from the
thalamus mainly terminate in
Brodmann area 3, although some
terminate in areas 1&2, of
somatic sensory area I (S-I).
There is redundant projection from
the thalamus to a poorly-understood
somatic sensory area II (S-II), which
also receives inputs from S-I. S-II is
somehow associated with exploratory movements.
Sensory input from the thalamus arrives primarily
in layer 4, although diffuse projections from lower brain centers
to layers 1&2 can activate whole regions at once. Descending projections
from layer 5 to the spinal cord and layer 6 to the thalamus provide direct
feedback control of sensory input.
|
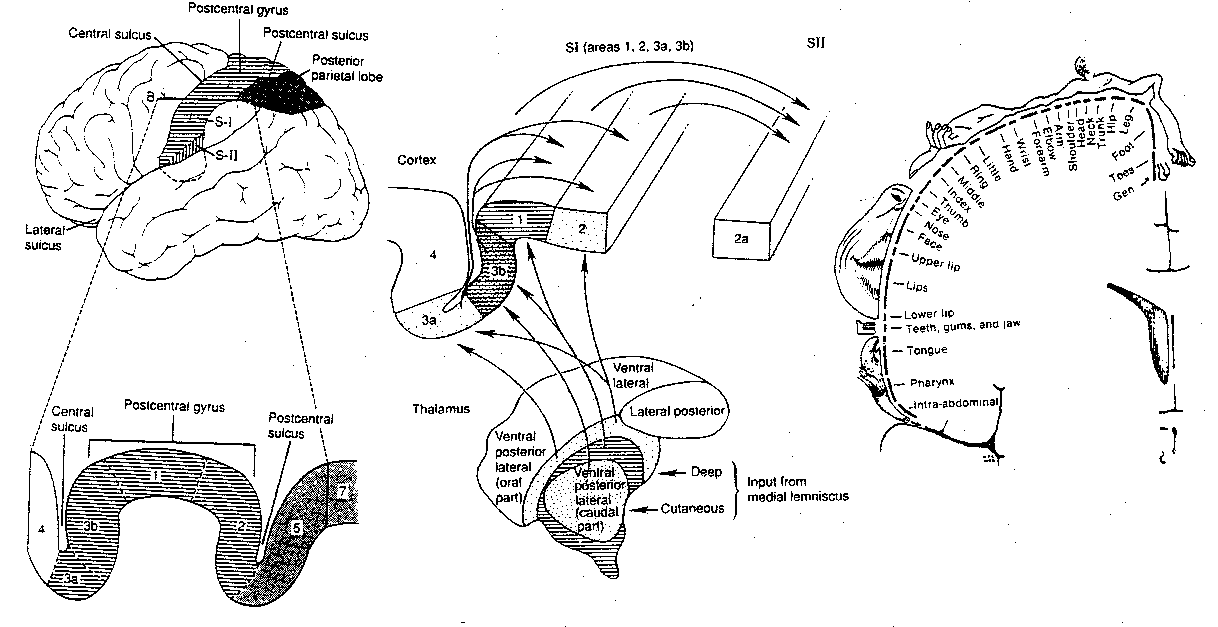
Brodmann area 3a & 2 receive a disproportionately large amount of
input from muscle, from tendon and from joint stretch receptors. Many of
these signals to 3a spread to area 4 of the adjacent motor cortex. Most
thalamic inputs go to area 3b, which receives inputs originating in the
cutaneous mechanoreceptors. Brodmann area 1 receives input from rapidly-adapting
cutaneous receptors concerned with texture, whereas area 2 receives input
from deep pressure receptors concerned with size&shape. Areas 3a, 3b, 1
and 2 all have topographical maps of the
body surface. The map for area 1
is shown in the illustration. Notice the small amount of cortical area
devoted to the arm, shoulder, neck & trunk — in contrast to the
wide areas devoted to hands, fingers, lips and tongue.
|
Digits of the hand of a monkey which are in frequent use will increase their cortical area. If the middle finger is removed from the hand of a monkey, within weeks the corresponding cortical area is taken-over by expansion of representation of the second and fourth finger. Cutting the sensory nerves from the arm of a monkey results in the arm's cortical area being taken-over by the lower face, although this process takes years.
Brodmann areas 5 and 7 are somatic association areas, which receive inputs from S-I and S-II. Electrical stimulation of these areas can cause a person to have an experience of feeling an object (such as a ball or knife).
There are 4 primary taste sensations: sweet, sour, salty and bitter. Just as the 3 primary colors can be used to produce a wide variety of combinations & shades, there can be many combinations & shades of tastes. Moreover, the perception of flavor often includes olfactory information. The standard substances used to index tastes are sucrose, hydrochloric acid, sodium chloride and quinine, respectively, all of which are given an index of one. By this standard, fructose has a sweetness index of 1.7 and caffeine has a bitterness index of 0.4 — meaning that a standard amount of fructose will be 1.7 times as sweet as the same amount of sucrose.
Taste impulses from the tongue&pharynx terminate on neurons in the gustatory nucleus of the solitary nuclear complex in the medulla. Impulses are then relayed to the ventral posterior medial nucleus of the thalamus, and then onward to the primary gustatory cortex in Brodmann area 3b (adjacent to the somatosensory area for the tongue). Another thalamic projection goes to a region in the insula (a region of the cerebral cortex buried in the lateral sulcus). Patients with high salt loss due to destruction of the cortex of the adrenal gland display an extraordinary craving for salty foods — although this mechanism is probably not primarily mediated by the cerebral cortex.
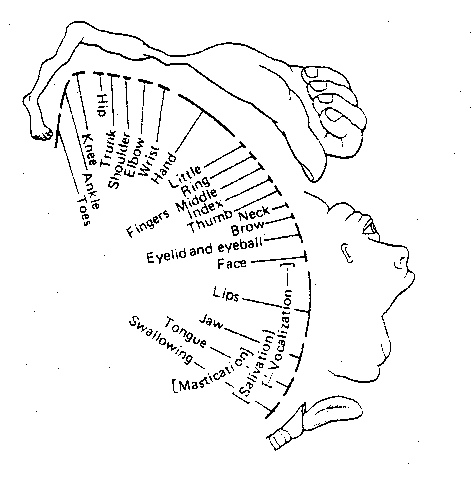
|
Brodmann area 4 is the primary motor
cortex (somatomotor cortex), although
the premotor "association" cortex in
area 6 also makes direct contributions
to voluntary movement. Neither area 4
nor area 6 possess a layer 4 (the
granular layer), and hence these areas
are called the agranular cortex.
Like the somatosensory cortex, these motor
areas also have topographical maps of the
body. The illustration shows the map of
Brodmann area 4. This map is similar to
area 1, except that an even greater area is
devoted to the hands and fingers. Also, the
somatosensory cortex allocates more area for
superficial structures of the face, whereas
the somatomotor cortex is more concerned with
structures responsible for mastication
& vocalization.
![[internal capsule]](./FigVI10.gif)
Nerve fibers from the motor areas
join fibers leading to&from other areas
of the cortex (the corona radiata) to
form a bundle of fibers called the
internal capsule, which adjoins the
basal ganglia. Some of these fibers from
the motor cortex terminate on the basal
ganglia as part of what is often referred
to as the extrapyramidal system. A large
portion of the internal capsule connects
the cerebral cortex to the spinal cord and,
hence, is known as the corticospinal tract.
![[corona radiata]](./FigVI11.gif)
![[pyramid on ventral brainstem]](./FigVI12.gif)
In the midbrain, this bundle of axons is known as the cerebral peduncle (or crus cerebri), and in the medulla it forms the pyramid. Hence, the corticospinal tract is also called the pyramidal tract. Fibers that synapse with the red nucleus in the midbrain, the nuclei of the pons and the inferior olivary nuclei of the medulla, all communicate with the cerebellum. Neurons in the red nucleus are most activated by exploratory (rather than routine) movements of the fingers.
About 30% each of the fibers in the pyramidal tract come from area 4
and area 6. The other 40% come from the somatosensory cortex, providing
feedback on sensory signals. 90% of the pyramidal fibers have a diameter
of less than 4 microns, but fibers originating from the giant Betz
pyramidal neurons (found only in area 4) have diameters of about 60 microns.
With conduction speeds of 70 meters per second, these are the most rapid
signals coming from the spinal cord.
![[SMA and PMC inputs and outputs]](./FigVI13.gif)
The posterior and lower portion of area 6 is known as the premotor cortex (PMC), whereas the anterior and upper portion of area 6 is known as the supplementary motor area (SMA). Although 80% of fibers in the pyramidal tract originate from the primary motor cortex (area 4), the remaining 20% originate from the PMC and SMA. Outputs from the SMA and PMC are mainly to area 4, but the PMC sends some output to the nuclei of the reticular formation. Nerve signals from the premotor area leading directly to the pyramidal tract might, for example, position the shoulders and arms so that the hands would be oriented for the performance of a specific task. The PMC is most strongly activated when voluntary movements are carried out with the guidance of sensory information, as (for example) when proceeding through a maze. The PMC is also particularly active during the performance of new, rather than routine, motor activities.
Memory of learned motor sequences (motor subroutines) seems to be stored in the SMA. Monkeys in whom the SMA has been lesioned show an awkward orientation of the hand when attempting to perform manual tasks. Tasks requiring the coordinated activity of two hands are also severely impaired. Human patients with SMA lesions initiate voluntary movements less often. Victims of SMA lesions begin to show signs of recovery within a few weeks, but recovery is never complete. Human subjects mentally rehearsing a sequence of finger movements without actually executing them, show increased blood flow in the SMA, but not in the other cortical motor areas. In sum, the SMA seems to be important for planning, initiating, remembering and executing motor sequences.
The premotor areas receive inputs from the prefrontal and parietal association areas. The Brodmann area 7 in the parietal association cortex integrates information from visual, auditory and somatosensory input to locate objects in space. A bundle of nerve fibers (the superior longitudinal fasciculus) communicates this information to area 6 to direct movement. The premotor areas also receive input from part of the limbic cortex (area 24), which is thought to contribute motivational input to motor planning.
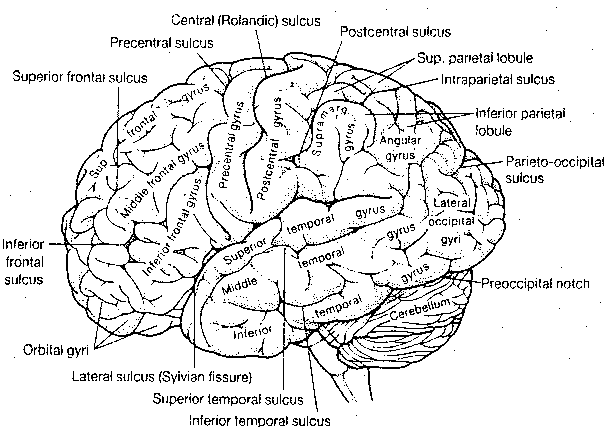
The subdivision of the cerebral
cortex into the frontal, temporal,
parietal and occipital lobes is based
on the names of the overlying bones in
the skull. Although this division is
artificial, it can be useful.
![[gross functional subdivisions of the cerebral cortex]](./FigVI15.gif)
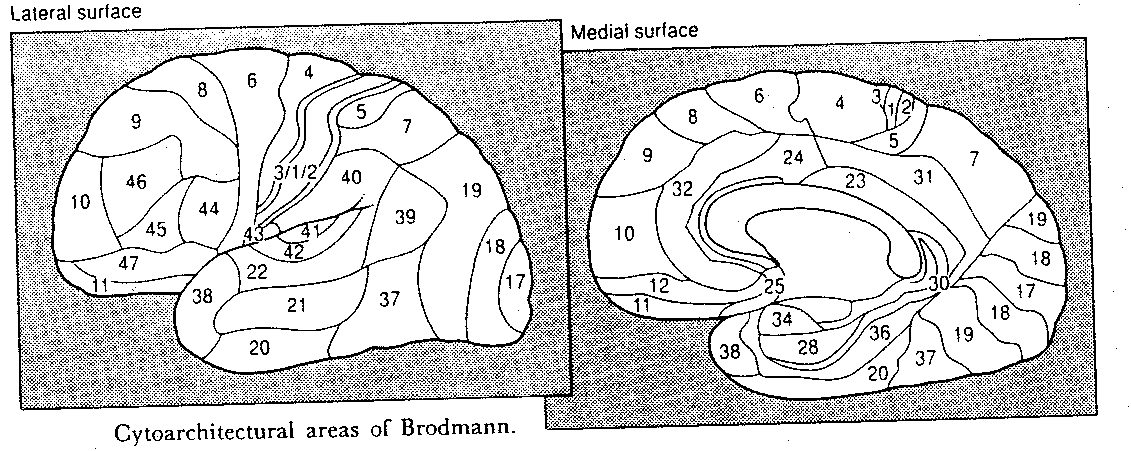
It should be clear from the foregoing material that the cerebral cortex contains several regions which are devoted to being receiving areas for special senses, and motor regions for voluntary control of muscular action. By default, cortical areas not specifically associated with sensory reception or motor dispatch are regarded as "association areas", even when the function of some of these areas is totally unknown. Since virtually all sensory areas are posterior to the central sulcus, areas not devoted to primary sensation in the temporal, parietal and occipital lobes are regarded as "sensory association cortex". Anterior to the central sulcus, in the frontal lobe, the association areas are deemed to be important for planning of motor activity. Portions of the cerebral cortex associated with the limbic system, such as the medial cortex and parahippocampal gyri, are often called the "limbic association cortex". These areas are presumably important for motivation and emotion.
Association cortex may well be the most subtle and complex brain tissue, and it is also extremely difficult to study. The association cortex of animal subjects is less comparable to human brains than sensory or motor cortex. Natural lesions to human brains, such as strokes, cancers or missile wounds, rarely cause damage with a high degree of specificity — and even more rarely occur of both sides of the brain in the same region.
The Electroencephalograph (EEG) uses electrodes placed on the
scalp to measure underlying electrical activity of the brain. EEG studies
have been disappointing in terms of what they have taught us about mental
function. A Magnetoencephalogram (MEG) is a magnetic counterpart to
an EEG, except that 3-dimensional localization is possible for the groups
of neurons generating magnetic fields — with isocontour maps corresponding
to field intensities. The MEG probe requires a Semiconducting QUantum
Interference Device (SQUID) which must be immersed in liquid helium to
maintain superconductivity.
![[CAT scans (= CT scans)]](./FigVI16.gif)
Computerized Axial Tomography scans ("CAT scans" or "CT scans") use an X-ray source coupled with an X-ray detector on the opposite side of the head, which are rotated together to give a cross-sectional picture of the brain at one level or cut. CT scans are of greatest value for showing physical changes in brain tissue, although small tumors may be missed if absorption properties are like those of normal tissue. Positron Emission Tomography (PET scans) differ from CT scans in that PET scans provide information about dynamic states of the brain, such as local blood flow or local glucose utilization. For example, glucose tagged with radioactive fluorine emits positrons (electrons with positive charge) which interact with electrons to produce gamma rays. Spatial resolution is not much better than one centimeter, although this could improve. The biggest drawback to PET scanning is the high cost, due to the need for a cyclotron to produce short-lived radioactive isotopes. Magnetic Resonance Imaging (MRI) takes advantage of the fact that the nuclei of certain atoms, hydrogen and phosphorous in particular, behave like a magnet. When those atoms are placed in a strong magnetic field, the nuclei align the axis of spin with the direction of the field. When the field is turned-off, the nuclei release a characteristic radiofrequency emission. MRI can detect the subtle differences in hydrogen concentration in different tissues, and the phosphorous of ATP is most concentrated in areas of high metabolic activity.
Brodmann area 18 is regarded as a secondary sensory area concerned with the elaboration and synthesis of visual information. Area 19 is well-connected with many other cortical regions, and thus is believed to be concerned with integrating visual information with information from the other senses. Most high-order processing of visual information seems to occur outside of the occipital lobe. Projections to area 39 in the parietal lobe are concerned with symbols, including numbers and letters. Projections to area 7 are concerned with movement and stereoscopic vision. Projections to area 20/21 in the temporal lobe are concerned with detail and color. Electrical stimulation of area 21 may produce visual hallucinations.
Some patients with lesions in area 19 on the left side lose their ability to read without losing their ability to write. Sometimes these patients can read individual letters and spell whole words without understanding them. Lesions in area 19 to fibers leading to the temporal lobe can result in a patient who can describe the features of an object without being able to say what the object is. Patients with lesions in area 37 can see faces without recognizing them, although they can describe the nose, cheeks, mouth, etc. These same patients can identify objects by class, but not by specific type. For example, the patient can say whether an object is a car, a bird or a tree, but cannot say whether it is a Volkswagen, an eagle or a pine. Lesions to both sides of the dorsal region of area 19, toward the parietal lobe, sometimes result in a patient who is unable to perceive a visual field as a whole, although individual objects can be identified. Such a patient may be unable to recognize the significance of playing card combinations in a "hand", although the individual cards can be identified. The patient may also have difficulty searching the visual field for an object.
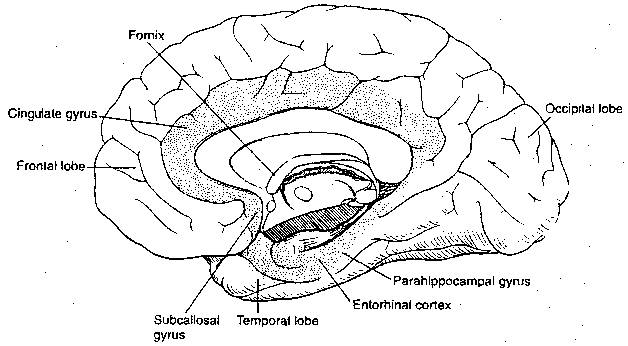
The cerebral lobes are not functional units, and this is particularly obvious with the temporal lobe. The medial and anterior portion includes the olfactory association cortex as well as areas considered to be "limbic association cortex". As has been mentioned, the lower portions of the temporal lobe, areas 20/21, are concerned with processing visual information. The upper portions of the temporal lobe are concerned with processing auditory information. Much of this area is only a part of a "language cortex" area that includes portions of the parietal and frontal lobes. The posterior third of area 22 is known as Wernicke's area and it, along with area 39 of the parietal lobe, is important for language understanding. Area 44 and part of area 45 in the frontal lobe, known as Broca's area is important for language production (spoken or written).
Broca's area is connected to Wernicke's area by a bundle of nerve fibers known as the arcuate fasciculus. Patients with lesions to the arcuate fasciculus show good comprehension of spoken language and reasonably fluent conversational speech, but have great difficulty repeating the spoken language they hear. They show difficulty reading aloud, despite good reading comprehension. The patient is often unable to name objects pointed-to, and writing typically shows errors in spelling and word-order.
PET scans of Broca's area show increased activation for tests generating lists of verbs associated with a noun (eg, "flower" would make one think of the verbs grow, cut, smell, etc.). Patients with lesions to Broca's area (Broca's aphasia) may be mute initially, but they usually recover, at least partially. Their speech is dominated by nouns and verbs. "Do you want to go to the store with me" might be expressed as "go store me". Although Broca's area is primarily concerned with language production rather than with comprehension, it does seem to play a role in the understanding of grammatical aspects of language. Patients with lesions in Broca's area often have difficulties with questions like "Is it true that my mother's brother's sister is a female?" Patients with Broca's aphasia also often have "tip-of-the-tongue" word-finding problems which can be offset by telling the patient the initial syllable or some other clue.
Damage to Wernicke's area (Wernicke's aphasia) is primarily
characterized by lack of comprehension of spoken language. If the lesion
extends into the parietal lobe, comprehension of written language may be
affected. Patients who comprehend written language can sometimes learn
to communicate through sign language. Often, however, the patient has no
concern or awareness that a problem exists. Wernicke patients speak in a
jumble of words that convey little meaning. For example, when one Wernicke
patient was asked how he was, he replied, "I felt worse because I can no
longer keep in mind from the mind of the minds to keep me from mind and up
to the ear which can be to find among ourselves". Although patients with
lesions to the arcuate fasciculus and to Broca's area often show
considerable recovery, recovery is poor for Wernicke's aphasia.
|
Area 40 (the supramarginal gyrus) of the parietal lobe is associated with the correlation of sounds and letters. Patients with lesions in this area may be unable to write the letters "B-A" upon hearing the spoken syllable "BA". Patients with lesions in area 39 (the angular gyrus) of the parietal lobe have difficulty writing words that are not phonetic, or which are phonetically ambiguous, for example "circuit" (which could be "surkit", for example). When reading aloud, these patients demonstrate "deep dyslexia", extracting word-meaning without translating written words into sounds, and saying "fowl" or "sofa" when having read "bird" or "couch". Lesions to the angular gyrus of Japanese readers impairs their ability to read Kana (syllabic), but not Kanji (ideographic) writing. PET scans of normal Japanese in the process of reading shows activation of areas 39&40 for Kana, and activation of areas 21&37 for Kanji.
There is some evidence that storage of memories of life-events occurs in the temporal lobe. Electrical stimulation of the cortex has evoked real or imagined experience of previous happenings only in the temporal lobe. However, this is almost always only seen in patients with temporal lobe epilepsy, and only 8% of temporal lobe epilepsy patients demonstrate an experiential response to electrical stimulation of the temporal lobe. Patients with temporal lobe epilepsy have the experience of "deja vu" more frequently than normal subjects, as well as the experience of "jamais vu" (lack of a feeling of familiarity with familiar stimulus patterns). During an attack of temporal lobe epilepsy everything may appear visually larger or smaller, and the patient has a feeling of unreality or of being detached from his own body. Many patients with temporal lobe epilepsy develop a personality syndrome characterized by a loss of interest in sex, intense emotionality and ardent religious interest. These patients can be socially aggressive, extremely moralistic and lacking in humor.
The importance of areas 5&7 for body orientation in space have already been discussed in connection with the somatosensory cortex. The importance of areas 39&40 for language have been discussed in connection with the temporal cortex. Patients with parietal lesions may have difficulties solving geometrical puzzles, especially ones that are 3-dimensional.
Patients with lesions to that angular gyrus of the left hemisphere have difficulty identifying and orienting their fingers. Such individuals also have difficulty with simple arithmetic, which led to the hypothesis that this is somehow related to the fact that children often learn to count with their fingers. Nonetheless, some patients who have no trouble with the subtraction "796-342" will fail repeatedly on "534-286", possibly because of the "spatial" requirement to "carry-over". These patients become confused by grammar involving spatial syntax, such as the difference between "my brother's wife" and "my wife's brother". And there can be great difficulty distinguishing left from right. A particularly troublesome task for such person is the request to "touch your left thumb to your right ear".
The prefrontal cortex, areas anterior and inferior to the SMA, constitutes nearly one-third of the cortical surface, yet it is very difficult to study and its functions are poorly understood. A gross subdivision is often made between the orbitofrontal cortex (areas 11&12) and the superolateral cortex (areas 9,10,45&46). The orbitofrontal cortex is often associated with ethical, social or emotionally related behavior. The superolateral cortex is often called the "executive centre" of the brain. A portion of this area which has proven amenable to animal studies corresponds to area 46, a locus of so-called "working memory".
A hungry monkey will be shown where a morsel of food is hidden. A barrier will temporarily obscure the monkey's view, and then when the barrier is removed the monkey will find the food. Damage to area 46 specifically interferes with the ability of a monkey to perform this task. Working memory represents a model of the world that persists in the mind in the absence of sensory input. Working memory allows for a dissociation between immediate stimulus and immediate response. It is working memory that allows a driver to move her hand from the steering wheel to the gearshift without taking her eyes off the road. Working memory is also called "scratch pad memory", because it can be used to store phone numbers in the seconds between look-up and keying. Working memory can also be referred-to as "active memory" because it may consist of activated memories retrieved from long-term storage.
The classic test for superolateral function in humans is the Wisconsin Card Sorting Test. In this test, subjects are asked to sort a deck of cards according to some criteria, eg, sort the cards by number. Patients with superolateral cortex lesions who have been sorting according to number, who are told to change the sorting criteria to sort-by-color will continue to sort by number. Such "perseveration" is an indication of the use of "associative memory" (rather than "working memory") to direct activity. Patients with superolateral lesions not only show "pathological inertia", but they are easily distractible (have a reduced capacity for attention). Tasks which require simultaneous retention of several facts cannot be performed.
At birth an infant's brain is one-fourth the weight of an adult brain, but by the age of 2 brain weight has tripled to be three-fourth's of adult brain-weight. Prior to eight months of age, a human infant is typically incapable of performing the working memory task of which an adult monkey is capable. A child's prefrontal cortex is not believed to be fully developed until puberty. A child's version of the card sorting test is the game "Simon Says". Working memory must retain the information to not follow a command if it is not given with the phrase "Simon Says". In one battery of tests, children's performance on Simon Says Tasks increased from between 25% to 75% between the ages of 4 and 6.
The executive role of the superolateral cortex is indicated by the fact that no other cortical region is so capable of initiating action independent of external stimuli. In this sense, it is said to regulate the temporal control of behavior, separating immediate movement from immediate sensation. It formulates plans, implements them, appraises the results and does error-correction in response to feedback. The superolateral cortex contains "metamemory", ie, knowledge of one's intellectual capabilities and knowledge. It contains managerial knowledge for "autopilot", allowing someone to drive a car and hold a conversation at the same time. It allows for planning that takes into account the probable future consequences of one's actions.
Patients with frontal lobe lesions on both sides of their brain often display no impairment on various intelligence tests, but family, friends and employers notice "personality changes" reflecting the cortical functions described. Such patients are passive, impulsive, and show poor judgement as well as lack of concern for the consequences of their actions. Certain intelligence tests do show severe decline in performance, especially those requiring multi-step analysis or abstract thinking. A frontal lobe patient may be unable to explain the meaning of the phrase "Don't cry over spilt milk".
The orbitofrontal cortex (areas 11&12) has been called the centre of
the "social self". Whereas patients with superolateral lesions show increased
passivity, patients with orbitofrontal lesions show disinhibition — often
judged to be a "lowering of ethical and moral
standards". Orbitofrontal lesion patients have a
decreased control of impulses, which often has disastrous social consequences.
These patients can be coarse, irritable, insulting, hypersexual and
hyperactive.
|
The so-called "limbic association" area is that part of the cerebral cortex included in the limbic system. The area includes portions of the parietal, temporal and frontal cortical lobes. The posterior portion of the orbitofrontal cortex is included in the limbic system, and the anterior cingulate ("belt-shaped") gyrus (area 24) is a part of the limbic association cortex which has extensive connections with the superolateral cortex. Intentional lesions to area 24 can reduce anxiety and obsessive-compulsive behavior, but such patients often become less meticulous and lose interest in many activities.
Prefrontal lobotomy (leucotomy, cutting of the white matter),
was first suggested as a means of treating mental illness by severing the
connections between the frontal and limbic cortex. By the late 1940s
many psychiatrists regarded leucotomy as the treatment of choice for patients
with a "fixed state of tortured self-concern", including hypochondria,
anxieties, worrying and "guild-laden ruminations about the past and fearful
anticipation of the future". Follow-up studies showed less severe long-term
consequences for orbitofrontal leucotomies than superolateral ones, but the
use of leucotomies declined sharply in the 1950s.
|
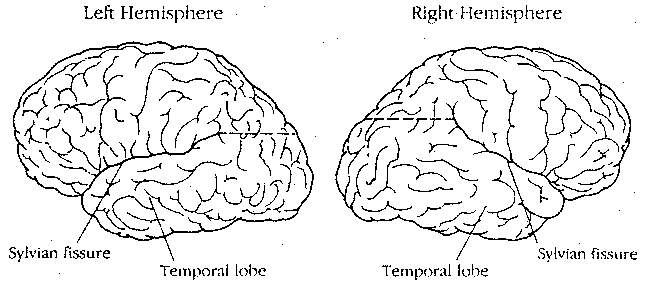
The cerebrum consists of a left hemisphere and a right hemisphere, which are anatomically unequal in most cases. For most right-handed individuals, the height of the posterior end of the Sylvian Fissure is higher on the right hemisphere, whereas this is only true for a minority of non-right handers. The right frontal lobe tends to be wider than the left, whereas the left occipital lobe tends to be wider than the right in right-handed individuals.
The left hemisphere receives sensation from and controls the right side of the body, and vice versa. About 90% of the population is classified as right-handed in all current races and cultures (Cro-Magnon peoples were evidently 80% left-handed). Some notable left-handers have been Alexander the Great, Leonardo Da Vinci, J.S.Bach and Albert Einstein. The English word derives from the Celtic lyft (meaning weak), the French word gauche means "awkward" and the Latin word sinister means "evil" — indicating a long history of cultural biases. A Johns Hopkins University study of mathematically gifted students found twice the expected proportion of left-handers.
The left hemisphere is dominant for language in 96% of right-handers and about 70% of left-handers. The language functions described earlier for Broca's area and Wernicke's area are found on the left hemisphere for about 93% of the total population, on the right hemisphere for about 5% and mixed in the rest. I will adopt the common practice of using the phrase "left-hemisphere" to refer to the "language-dominant hemisphere" for the sake of clarity and simplicity.
In fact, Broca's (area 44) and Wernicke's (posterior of area 22) areas on the right hemisphere are "dominant" for different language functions than those on the left. Patients with damage to area 44 on the right hemisphere speak with flattened intonation and have difficulty judging the emotional tone in the speech of others. Such patients often make statements such as "I am angry and I mean it" to compensate for their failure to convey emotion through tone of voice. PET scan studies of professional musicians shows activation of area 22 on both hemispheres when they listen to musical pieces, but only on the left hemisphere when they listen to musical scales. (The composer Maurice Ravel developed Wernicke's aphasia and could no longer write music, despite the fact that he could still discern the smallest mistakes when listening to a musical performance.)
Patients with damage to the left parietal lobe are unable to brush
their teeth, strike a match, wave goodbye or perform any number of other
voluntary actions when asked to do so, despite retaining the capacity to
do these actions spontaneously in appropriate situations. Patients with
damage to the right parietal lobe seem to lose all consciousness of the entire
left hemisphere. Only the right side of the face is shaven, only food on the
right side of the plate gets eaten, and when asked to draw a clock, the patient
will only show the numbers on the right side.
![[drawings by singly cerebral-hemisphere-injured patients]](./FigVI23.gif)
Many sweeping generalizations
have been made about right and
left hemisphere function which
are hard to justify. One such
claim is that the left
hemisphere is specialized
for local, detailed,
serial processing, whereas
the right hemisphere is more
specialized for global,
holistic, parallel processing.
Some evidence for this comes
from patients who draw
pictures from recall. Patients
with right-hemisphere damage
recall details while missing
global features, whereas
patients with left-hemisphere
damage do the reverse.
Left-damaged patients have
difficulty with verbal
material, whereas right-damaged
patients have difficulty with
spatial problems. Left-damaged
patients tend to display
feelings of anger&despair,
whereas right-damaged
patients show placidity
and sometimes even elation.
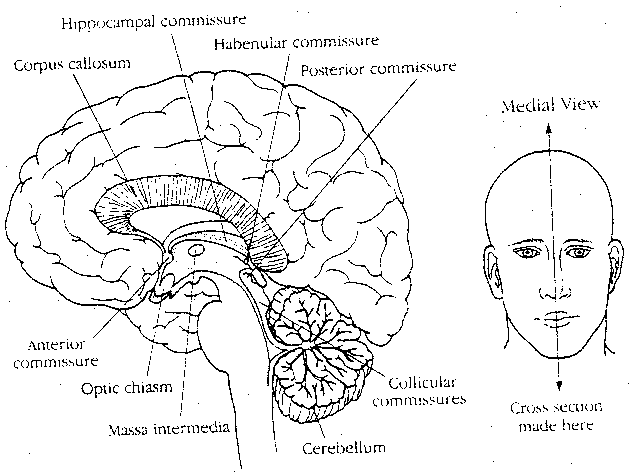
|
A number of commissures contain fibers which connect the two hemispheres, but only two contain fibers connecting the cerebral hemispheres: the anterior commissure and the corpus callosum. The anterior commissure contains some fibers interconnecting the olfactory bulbs, but the great majority of fibers connect regions of the inferior and middle temporal gyri. The corpus callosum connects neurons in the association (not sensory or motor) cortices of each side.
Patients in whom the corpus callosum has been cut (for the treatment of epilepsy), can easily name objects presented to their right visual field, but deny awareness of objects presented to their left visual field. If, however, the letters D-O-G are presented to the left visual field, the patient can use his left hand to select a model of a dog from amongst a group of models. The left hand/left visual field/right hemisphere and the right hand/right visual field/left hemisphere appear to be systems entirely divorced from one another. Although the right hemisphere can process very simple visual inputs, it is not capable of language output, and can only express itself through actions of the left hand. When the corpus callosum and optic chiasm of a cat is cut, if the cat is trained to do a task with a patch over one eye, it will require complete re-training to do the same task when the patch is moved to the other eye.
The most dramatic outcome of callosotomy, however, is the alien hand sign. In one case history, the clinician writes: "...she would be putting on clothes with her right hand and pulling them off with her left, opening a door or a drawer with her right hand and simultaneously pushing it shut with the left." Another clinician describes cases wherein the patient's "left hand would not allow him to smoke and would pluck lit cigarettes from his mouth, whereas another patient's left hand (right brain) preferred different foods and even different television shows and would interfere with the choices made by the right hand (left hemisphere)". Evidence indicates that the corpus callosum not only functions for the exchange of information between hemispheres, but for one hemisphere to assert dominance over the other hemisphere.
The question of whether cutting the corpus callosum produces two independent minds has not been resolved, but it could have interesting implications for cryonics. One can imagine a person making independent cryopreservation arrangements with different cryonics organizations for each half of his head on the theory that it acts as an insurance against one of the organizations failing. Under ideal circumstances, the two halves would eventually be re-united in reanimation, but one could imagine circumstances in which one or both of the halves serves as a template for reconstruction of a whole new individual. What becomes of the original identity in such cases?
![[gross subdivisions of the cerebral cortex]](./FigVI14.gif)
![[projections from the visual cortex]](./FigVI17.gif)
![[temporal association cortex projections]](./FigVI18.gif)
![[working memory experiment in a monkey]](./FigVI20.gif)
![[GO TO BEN BEST'S HOME PAGE]](../../homeback.gif) HOME PAGE
HOME PAGE